














�������C���ɏ���Ă����A���͂ǂ��i�����Č��E���L�����̂��B���z���E���̐��藧���ƍ���̕ω���?
|
�w����N���C���U�z�A���ɂ����镪�z���E�̌���v���̉𖾁x ����29��i�ߘa2�N�x�j�`��30��i�ߘa3�N�x�j������ ���s��w ��w�@���w������ �A���w���� �A���n�����ފw������(���l�������w) �y���� ���R �_�i���� �i���w���m�j 1978�N�A�����s���܂�B2001�N�A�����s����w���w�����ƁB03�N�A�����w�@���w�����ȏC��(���w�C�m)�B06�N�A������w��w�@���w�n�����ȏC��(���w���m)�B��t��w���w�������w�� �w�p�����x�����B07�N�A�����w�@���w������ JSPS���ʌ�����PD�B10�N�A����w�@ ���C�������B�E�B�[����w�A���n���i���w���� JSPS�C�O���ʌ������B12�N�A���呍������������ ���C�����B15�N�A�ӂ��̂��ɒn�����j�~���[�W�A�� �y�����B18�N�`���݁A���s��w��w�@���w�����Ȑ����Ȋw��U �y�����A���������٘A�g�����B���͐A���n���i���w�A�����n���w�A���א����w�𒆐S�Ƃ������l�������w�B���l�ȗ���A�����ǂ̂悤�ɐi�����āA���݂̕��z����l������Ɏ������̂��𖾂炩�ɂ��Ă����B�l�X�Ȓn���A����ΏۂɃt�B�[���h���[�N���烉�{���[�N�܂ŕ��L���A�n���i����W�c�\���̌��n����푽�l���̈ێ��E�n�o�@�\�̗�����ڎw���B��� : 09�N���{�A�����ފw���܁A16�N���{�A���w���܁B�_�������B |
|


�u�n�}�I���g���v�̓�ƐA�����z���E�̉𖾂ɒ���
| ---- �A���̂������E�������n�߂�ꂽ�������������������������B | |||
���w���̍��͍����̏W���D���ŁA����������߂܂��邱�Ƃɋ���������܂����B���������炵�Ă����Ȃ��A���w���炢���琶������ɔ��R�Ƃ����S������܂����B�N���m��Ȃ����Ƃ��𖾂������Ƃ����v���������Ȃ�A���Z�ł̏����̖��͌����҂ł����B�����āA�s����̗��w�������w�Ȃɐi�w���A�w�ƂƑ������̕������ɖv���B�w��3�N���̖�O���K�I�����A�������̑������̌��ˍ����ŁA�B�ꗚ�C�\�������A���n�����ފw�������̖�O���K�����܂����B����܂łǂ��炩�Ƃ����Ɠ����ɋ���������܂������A�̏W�����A�����K�������}�ӂƈ�v���Ȃ����Ƃɔ[���ł����A���ꂪ�A���w�ɂ̂߂荞�ނ������ƂȂ�܂����B�s����ɂ͓��{�̐A���w�̕��E�q��x���Y���m�̕W�{�𑽐����������q��W�{�ق�����A���ƌ������Ƃ��ď��}�������̐A���������n�߂܂����B���}���ŗL�̃����e���{�N�̓��Ԃł̌`�ԓI���`�I������˂��~�߁A��q�C���U�z�\�͂̑r�����B�����e���{�N�̋N�����𖾂��邽�߁A���m�ے��͓��召�ΐ�A�����ŁA�ĔM�ъC���U�z�A���̌n���n���w�I�Ȍ������J�n���܂����B
|
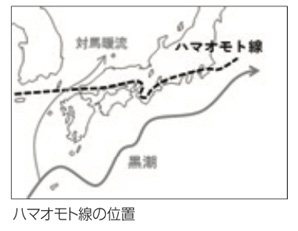

| ----�u�n�}�I���g���v�Ɠ���̊C���U�z�A���ɒ��ڂ��ꂽ�_�A���̒����̂���J��? | |
���m�ے��Ō��������ĔM�ъC���U�z�A���̃I�I�n�}�{�E�̕��z�̎��Ӓn��ł́A�ŗL�킪�핪�����Ă��܂��B���̒��ʼn��ш�܂ŕ��z���L�����̂��n�}�{�E�ł��B�n�}�{�E�ׂ�A�M�ыN���̊C���U�z�A�������z��M�шȊO�ɍL����ۂ̐i���I�v���Z�X��������Ǝv���܂����B���{�̓���N���̐A���A�n�}�{�E�A�n�}�i�^�}���A�n�}�I���g�A�O���o�C�q���K�I�A�n�}�A�Y�L�A�C���^�C�Q�L�A�����̕��z�̓n�}�I���g���Ƃ��������̕��z���E���Ƃقڈ�v���Ă��܂��B�n�}�I���g���͋���A���w�����̑��y�E���������搶�����A�N�ԕ��ϋC��15�x�A�~�̍Œ�C���̕���-3.5�x�̃��C���ł��B���̋��E���̌`���ߒ�������I�Ȏ�@�ŒT�邱�Ƃ��A��p�����҂̎g���Ƃ������܂����B�܂��A�n�����g���������̕��z�ɋy�ڂ��e���ɂ��Ēm���ł��A�n�}�I���g���͒��ڂɒl����ƍl���܂����B�n�}�{�E�̌�����{�i�I�Ɏn�߂��̂�2013�N�ŁA����10�N�߂������Ă��܂��B�t�B�[���h���[�N�͋�J�̘A���ŁA���{�ɂ͎��R�C�l���ƂĂ����Ȃ����߁A�A���������o���Ȃ����Ƃ���������܂��B���݂́A�������̊w�����ꏏ�Ƀe�[�}�Ɏ��g��ł��܂��B����ꂽ�f�[�^����A�ǂ�ȃX�g�[���[���a��������̂�����̃X�g�[���[������ɋ�������ɂ͂ǂ�������ǂ��̂����X�c�_��ςݏd�˂Ă��܂��B
|

���z���E�̌���v���A��`�I�ȈႢ���q���V��
| ---- �W�c�Q�m�~�N�X�E��q�U�z�\�́E�C���U�z�p�^�[���A���z���E�̌���v���Ƃ�? | |
���݂̕��z���悭�����A�����m�̈�`�\�����W�c�Q�m�~�N�X�̎�@�Ŕ�r���A�܂����ʐ��ƌʐ�����ʂ��܂��B���ʃp�^�[��������A�����ɂ͒n�j�I�Ȕw�i������͂��B����A�ʃp�^�[���͌ŗL�̗��j����ԓI�ȓ����ɋN�����Ă���\���������B���ɒ��ڂ��Ă���ŗL�̐��ԓI�����́A��q�̎U�z�\�͂ł��B�C���U�z�A���́A�A���킲�ƂɎU�z�\��(���V�ł��鎞��)�ɍ������邱�Ƃ��m���߂��Ă��܂��B���̎U�z�\�͂̍������z���E�̌���v���ɐ[���ւ���Ă���ƍl���Ă��܂��B�Ⴆ�A���݂ł����z���k�サ�����Ă����ł́A������瑊�����̎�q�̋������s���Ă���B���̏ꍇ�A�n���I�����ɑ��ւ�����`�\���͂ł��ɂ����B����A���ɒx�����x�ŕ��z���k��A�܂��͋��R�̎U�z�Ŗk�オ�N����A���z���`�����ꂽ��ł́A�e�n�W�c�̈�`�I�������i��ł����B�����A���ۂ̎�q�U�z�̗��j�͂킩��Ȃ��̂ŁA�t�Ɉ�`�\��������j��R�������Ƃ����̂����̌����ł��B�ŋ߂ł́A�C�����ǂ����V�����^�Ԃ��A�V�~�����[�V�������ł���悤�ɂȂ�A���V���̃X�^�[�g�n�_�ƗʁA���V���Ԃ�^���邱�ƂŁA����n��ւ̕��V���̓����m�������߂��܂��B������C���U�z�A���ɂ��K�p���A�C���̗���ƕ��z�̊W���q�̒蒅�ׂ����ƍl���Ă��܂��B
|

| ---- �C���^�C�Q�L�̈�`�\���ƁA�n�}�{�E�̌́E�n��ʂ̊C�����V���Ƃ�? | |||
�܂��A�C���^�C�Q�L�ɂ́A�u�엮���v�u�������v�u�k�����`��B��l����{�B(�k�����Ȗk)�v�̈�`�I�ɈقȂ�3�W�c�����邱�Ƃ�˂��~�߂܂����B���̌`���ߒ��́A��W�c�Ɩk�W�c�̕�������2���N�O�̍ŏI�X���̍��ŁA���݂̓��{�ɂ���C���^�C�Q�L�̂قƂ�ǂ͓엮���ȓ�ɓ������Ă����Ɛ�������܂��B�����������Ȗk�ɂ��ꕔ���c���ꂽ�W�c���������悤�ł��B�ŏI�X���ȍ~�A�C���g�ɂȂ�A�엮���ȓ�ƒ������Ȗk�̏W�c���k����͂��߁A�������ł͓엮���ȓ�Ɩk���̏W�c���𗬁B��������悤�ȁA��`�I�ɈقȂ�W�c������������ԂɂȂ����ƍl���Ă��܂��B�n�}�{�E�ł́A�����m���Ɠ��V�i�C���ő����̕�����������̂́A�e�n��ł͈�`�I�𗬂��p�ɂɂ���Ǝv���܂��B�����̌��ʁA�n�}�{�E��2�N�ȏ����q�����V�\���Ƃ킩��܂����B�����A��C���Ɏ�q���o�Ă����Ă��قƂ�ǂ͒蒅�ł��܂���B�������߂ɂ́A��q�����ɋ���邱�Ƃ��K�v�ŁA���̕��A��q�ɉh�{���̒��������Ȃ��Ȃ�A���V�\�͂ƒ蒅�\�͂̊Ԃɂ̓g���[�h�I�t�����݂���̂ł͂Ȃ����Ɨ\�z���Ă��܂��B��̏W�c�͊C���ɏ���Ăǂ����Œ蒅�ł���m���͍������A�k�̏W�c�ł͖��ʂȎU�z����������B�܂�A�k�̏W�c�ł͋ߏ�ł̔���\�͂ɓ������������A�����c��\���������킯�ł��B���ۂɁA��̏W�c�Ɣ�ׂ�ƁA�k�̏W�c�ŕ��V�����Ⴂ�W�c���������Ă��܂��B
|
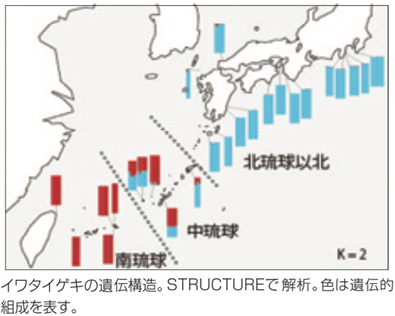

�V���Ȏ�ւ̃A�v���[�`�A���g���ł̕��z�ω���ǂ�
| ---- ����̂�������V�e�[�}������������肦�܂���? | |
����܂ŁA�n�}�{�E�ƃC���^�C�Q�L�̉�͂�i�߁A�قȂ��`�\���̑��݂�������܂����B�n�}�i�^�}�������l�̉�͂�i�߁A��r�������ƍl���Ă��܂��B�܂��A�������̊w���̓n�}�I���g�̌������i�߂Ă��܂��B����N���̐A�����A���ۂɂ͂ǂ�Ȍ`�����i�����邱�ƂŁA���{�ɂ܂Ő�����L���邱�Ƃ��ł����̂��B�����āA�n���̉��g������ω��̂Ȃ��A����ǂ̂悤�ɕ��z���ω����Ă����̂��A���ׂĂ��������ƍl���Ă��܂��B
�@�i��ޓ� �ߘa4�N5��31�� ���s�s����s��w���w���j |

